Welcome to the Buchberger group
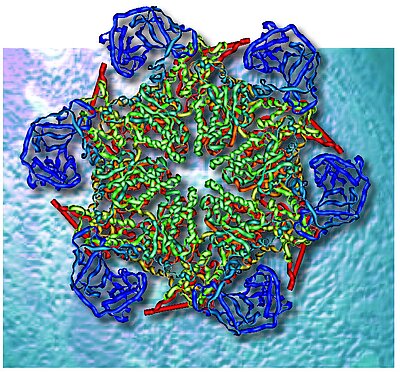
Our group is interested in the regulated intracellular degradation of proteins by the ubiquitin-proteasome system. The focus of our research is on the ubiquitin-selective, chaperone-related ATPase Cdc48/p97 - see above for an artistic representation. We use biochemical, cellular, and structural approaches to investigate mechanisms underlying target protein recognition and turnover. Our aim is to understand the pathological consequences of defective protein degradation in neurodegenerative diseases and cancer.
Selected Publications
Körner M, Müller P, Das H, Kraus F, Pfeuffer T, Spielhaupter S, Oeljeklaus S, Schülein-Völk C, Harper JW, Warscheid B and Buchberger A (2025). p97/VCP is required for piecemeal autophagy of aggresomes. Nature Communications 16:4243.
Körner M, Meyer SR, Marincola G, Kern MJ, Grimm C, Schuelein-Voelk C, Fischer U, Hofmann K and Buchberger A (2023). The FAM104 proteins VCF1/2 promote the nuclear localization of p97/VCP. Elife 12:e92409.
Tolay N and Buchberger A (2021). Comparative profiling of stress granule clearance reveals differential contributions of the ubiquitin system. Life Sci Alliance 4:.
Turakhiya A, Meyer SR, Marincola G, Böhm S, Vanselow JT, Schlosser A, Hofmann K and Buchberger A (2018). ZFAND1 recruits p97 and the 26S proteasome to promote the clearance of arsenite-induced stress granules. Mol Cell 70:906-919. Recommended by Faculty of 1000.
Böhm S and Buchberger A (2013). The budding yeast Cdc48-Shp1 complex promotes cell cycle progression by positive regulation of Protein Phosphatase 1 (Glc7). PLoS One 8:e56486.
Fernández-Sáiz V and Buchberger A (2010). Imbalances in p97 co-factor interactions in human proteinopathy. EMBO Rep 11:479-485.
Schuberth C and Buchberger A (2005). Membrane-bound Ubx2 recruits Cdc48 to ubiquitin ligases and their substrates to ensure efficient ER-associated degradation. Nat Cell Biol 7:999-1006.
For the full list of publications, please click here
Background
Intracellular Proteolysis
Regulated proteolysis plays a central role in numerous cellular processes, including protein quality control, cell division, signal transduction, DNA repair and many others. Eukaryotic cells possess two major proteolytic systems: the 26S proteasome and the lysosomal compartment (Fig. 1). Most soluble proteins are degraded by the 26S proteasome, a large, barrel-shaped protease complex consisting of four stacked rings. Its active sites are deeply buried in a central chamber formed by the two inner rings, and substrate proteins have to pass a channel gated by the distal rings in order to be degraded. The lysosome is an acidic organelle harbouring various proteases and other hydrolases. It is the end-point of two proteolytic pathways delivering membrane-confined cargo: the endolysosomal pathway targeting predominantly plasma membrane-derived proteins, and autophagy for the degradation of cytosolic content including some soluble proteins, insoluble protein aggregates, and even entire organelles, viruses and bacteria.

Ubiquitin
All three major routes for regulated proteolysis in eukaryotic cells are controlled by the covalent labeling of target proteins with ubiquitin, a small, highly conserved protein of 76 amino acid residues. This process (termed “ubiquitylation”, “ubiquitination” or “ubiquitinylation”) involves the formation of an isopeptide bond between the carboxy terminus of ubiquitin and the ε amino group of lysine residues on target proteins (Fig. 2). Ubiquitylation requires the energy of ATP and a catalytic cascade of three enzymatic activities E1, E2, and E3. The E1 (ubiquitin activating enzyme) activates ubiquitin by C-terminal adenylation, followed by formation of an energy-rich thioester with the active site cysteine residue. Activated ubiquitin is then transferred in a transesterification reaction to the E2 (ubiquitin conjugating enzyme), followed by modification of target lysine residues mediated by the E3 (ubiquitin protein ligase). The substrate specificity of the system is primarily determined by the large number (dozens to hundreds) of different E3 ligases found in all eukaryotes.
Ubiquitin itself can be ubiquitylated on either one of seven lysine residues and on its amino terminus, resulting in target protein modification by ubiquitin chains of different length, linkage type and complexity. In addition, ubiquitin chains are subject to editing by deubiquitylating enzymes (DUBs), further increasing the plasticity of the ubiquitin signal. Importantly, different ubiquitin chains mark substrates for distinct downstream processes (Fig. 2). For example, chains linked via lysine residue 48 of ubiquitin (“K48-linked chains”) are prototypical signals for targeting to, and degradation by, the 26S proteasome. By contrast, K63-linked chains, “linear” chains linked via the amino terminus of ubiquitin, and single ubiquitin moieties (“mono-ubiquitylation”) constitute signals for non-proteasomal fates of substrate proteins, for example in sorting processes of the endolysosomal pathway, signal transduction, and DNA damage repair.

Cdc48 / p97
The evolutionary conserved protein Cdc48 (also known as p97 and VCP in vertebrates and as TER94 in Drosophila) has emerged as an important motor and regulator for the proteasomal degradation of many proteins. Cdc48 is a chaperone-related member of the "ATPase associated with various cellular activities" (AAA) protein family. It converts chemical energy released by ATP hydrolysis into mechanical force believed to drive the “segregation” of ubiquitylated substrate proteins from stable protein complexes, membranes and chromatin, thereby facilitating their delivery to and degradation by the 26S proteasome.
Interestingly, Cdc48 has more recently also been shown to be critically involved in autophagy and endolysosomal protein degradation. Even though functional insights into the role of Cdc48 in the latter pathways are just beginning to emerge, it is intriguing to note that Cdc48 functions in all three major routes of intracellular proteolysis in eukaryotes. In line with such a central role in protein degradation, Cdc48 is essential in all organisms tested, and mutations in the human VCP gene cause neurodegenerative, muscular and motor neuron diseases.