Stomata & Guard Cell Action
Stomatal movement – guard cell physiology
In order to survive even extreme conditions, such as dry periods and/or heat, plants have to adapt their water balance to the particular environmental situations. Most important is thus the control of the water loss during the gas exchange via the stomata. The opening state of these microvalves is adjusted by special motor cells - the guard cells. This hydraulic motor reacts to internal as well as to environmental factors, which are detected by the guard cells via a multisensory network. These signals are then converted into the respective, situation-adapted stomatal movement. My workgroup played a decisive role in the elucidation of the guard cell engine components, consisting of ion channels, pumps and their control circuits. Thus, e.g. the effect of the drought stress hormone abscisic acid (ABA) is almost completely understood. Furthermore, we have already shown that guard cells can autonomously induce stomatal closure due to a sharp drop in air humidity.
Stomal opening width is crucial for CO2 absorption and water loss via the leaves. Therefore, an optimized water balance under heat and drought stress is dependent on the reaction of the stomata to the particular stress situation.
To open stomata a blue light receptor, together with effectors such as the 14-3-3 protein, acts on a proton ATPase, which, by generating a proton gradient, energizes the cell membrane. Following this driving force, potassium ions move into the guard cells via a channel, against their concentration gradient. At the same time, anions flow into the guard cell via a proton symport system, water follows the osmotic gradient, increasing the cell volume and opening the stomata (Figure 1). In principle, the ABA-induced stomatal closure runs in the opposite direction, but completely different systems are involved. ABA is bound by a receptor, which, in this activated state, inactivates a phosphatase of the PP2C type. The latter activates kinases (OST1 and calcium-dependent CPKs), which in turn activate anion channels by phosphorylation. The subsequent anion efflux depolarizes the cell membrane, thereby activating the potassium release channel GORK. As a result, water flows out, the cell volume decreases, and the stomata close (Figure 1). Both processes are regulated at the protein and the transcriptonal levels.
Investigation of stomatal movement is one of the most important branches of my research and is therefore reflected in a couple of projects.

ABA action and receptors
We described the primacy of abscisic acid (ABA) in the stomatal response to drying air and showed that guard cells possess the entire ABA biosynthesis pathway and that it appears up-regulated by positive feedback by ABA. When wild type Arabidopsis and the ABA deficient mutant aba3-1 were exposed to reductions in relative air humidity (rh) the aba3-1 mutant wilted while the wt did not. However, when aba3-1 plants, in which ABA synthesis had been specifically rescued in guard cells, were challenged with dry air they did not wilt (Figure 1). These data indicated that guard cell autonomous ABA synthesis is required for and is sufficient for stomatal closure in response to low rh. Guard cell autonomous ABA synthesis allows the plant to tailor leaf gas exchange exquisitely to suit the prevailing environmental conditions. A key element in ABA downstream signaling is the family of ABA receptors, the PYR/PYLs. In our current studies we are about to show that these receptors are not only involved in direct drought stress hormone ABA signaling but play also a role in the responses to other signals that lead to stomatal closure. Amongst other things, we therefore use a comprehensive microarray-based transcription analysis.

KlimaFit – Strategies for crop adaptation to the climate change
According to independent climatic models, in future a redistribution of annual precipitation and a rise in average temperatures are expected. The predicted climatic changes will affect crops in Bavaria in two ways. They have to survive under unfavorable water as well as temperature condition, which will result in significant crop shortfalls. Reliable scientific data on the response of crops to the anticipated stress combinations do not exist. Barley is one of the most important crops grown in Bavaria. The breeding selection of varieties, which can withstand both drought and heat, is tedious and difficult.
With our project, we want to identify genetic markers in barley, which might help breeders to identify tolerant strains. Thereby we are mainly examining drought stress. Here, the water management of the plants, which is controlled by pores on the leaf surface, the stomata, plays the most important role. The stoma opening width, regulated by the turgor of the two involved guard cells, is decisive for water loss through the leaves. Therefore, an optimized water balance under heat and drought stress is dependent on the reaction of the stomata to the particular stress situation. In this scenario, the stress hormone abscisic acid (ABA) plays a key role. As with all grasses, the barrier cell complex consists of a pair of guard cells neighbored by a pair of subsidiary cell (Figure 1). Stomatal closure, a first macroscopic response to ABA, is much faster in barley than in non-grasses and the dependent of the interaction of the guard cells (GC) with their subsidiary cells (SC). The nature of this cell-to-cell communication is so far almost unknown. Between both cell types, a highly regulated traffic exist in relation to signals and osmotically effective metabolites and ions. Thus, more insight into the communication between these two cell types is mandatory for the identification of potential barley marker genes and is the focus of our investigations.
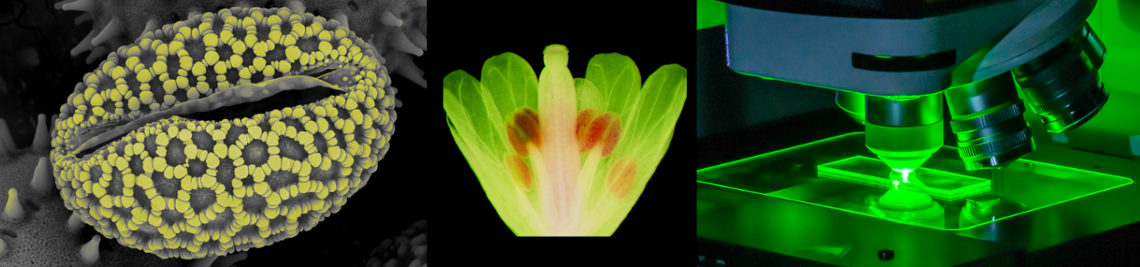
 and subsidiary (SC) cells. Fluorescein diacetate staining in transmitted light (A, C) and fluorescence images (B, D). (A, B) Both the GCs (yellow) and the secondary SZs (green) are intact in epidermal peels. (C, D) Mixing with a blender results in destruction of the (soft) SC and only the SZ survive.")