Start
Research
Groups
janzen lab
kramer lab
Topics
mRNA export
mRNA decay
mRNA Granules
SUPERRESOLUTION
MEMBRANE BIOPHYSICS
ADHESION
CYTOSKELETON AND MOTORS
QUORUM SENSING
TSETSE BIOLOGY AND PHYSICS
Telomere structure & function (THIS PROJECT WAS DISCONTINUED IN 2024)
Novel virulence factors in Leishmania parasites
Histone Methylation & Replication Initiation
NUCLEAR STRUCTURE AND DYNAMICS
HOST SKIN INFECTION
AFRICAN SLEEPING SICKNESS
PARASITE MOTILITY
ANTIGENIC VARIATION
THEODOR BOVERI
A Virtual Boveri Library
Cell Studies
Figures from Boveri Publications
TRAFFICKING OF GPI-PROTEINS
DZVT
Lecture Series
Team
Staff scientists
Christian Janzen
Nicola Jones
Nicola Jones - graduates
Nicola Jones - full list of publications
AG Beneke
Susanne Kramer
Publications
Manfred Alsheimer
Manfred Alsheimer - publications
Markus Engstler
Graduate students
Johanna Odenwald
Nicolas Hagedorn
Nicole Herrmann May
Narges Jamshidi Khameneh
Nashmil Aslam
Carina Praisler
Alexia Achilles Amaral
Melina Mitnacht
Kevin Bongers
Adamu Musa Muhammed
Undergraduate students
Technicians and IT
Silke Braune
Patrick Haag
Reinhild Fischer
Kathrin Weißenberg
Elina Krüger
Lotta Janner
Lidia Schegelski
Elisabeth Meyer-Natus
Yeva Dzhokych
Alumni
History
Teaching
Bachelor
Master
Lehramt
Medizin
Contact
Intranet
Likes
Deutsch
DE
Start
Research
Groups
Topics
Lecture Series
Team
Staff scientists
Graduate students
Undergraduate students
Technicians and IT
Alumni
History
Teaching
Bachelor
Master
Lehramt
Medizin
Contact
Intranet
Likes
Stop animation
Lehrstuhl für Zell- und Entwicklungsbiologie
Biocenter
Lehrstuhl für Zell- und Entwicklungsbiologie
Research
Topics
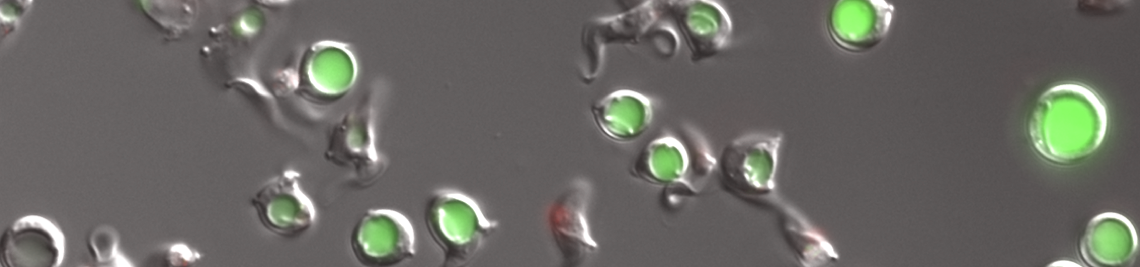
CYTOSKELETON AND MOTORS
CYTOSKELETON AND MOTORS
How to Find Us
Hubland Süd, Geb. B1
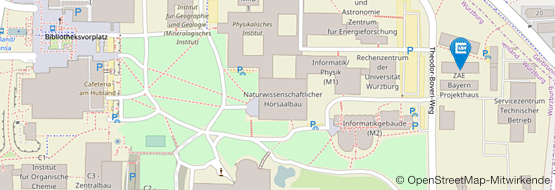
Hubland Nord, Geb. 32
Julius-von-Sachs-Platz 2
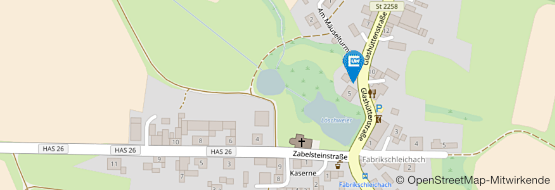
Fabrikschleichach
Hubland Süd, Geb. B2
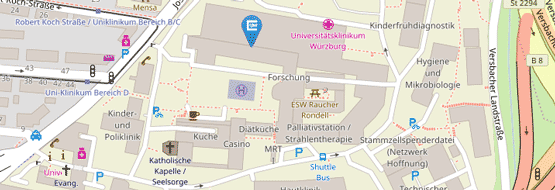
Campus Medizin
Hubland Süd, Geb. B1
Hubland Nord, Geb. 32
Julius-von-Sachs-Platz 2
Fabrikschleichach
Hubland Süd, Geb. B2
Campus Medizin
Picture credits
Privacy Disclaimer
Barrierefreiheit