mRNA Granules
mRNA Granules (Kramer lab)
Function and regulation of RNA granules
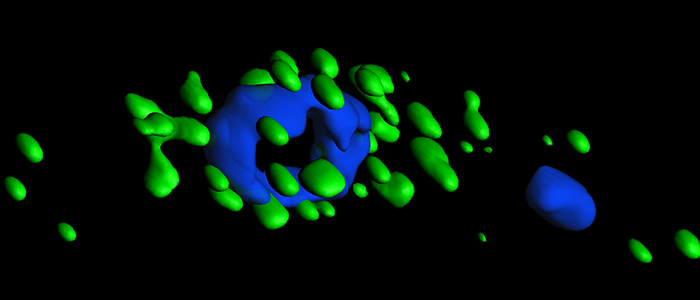
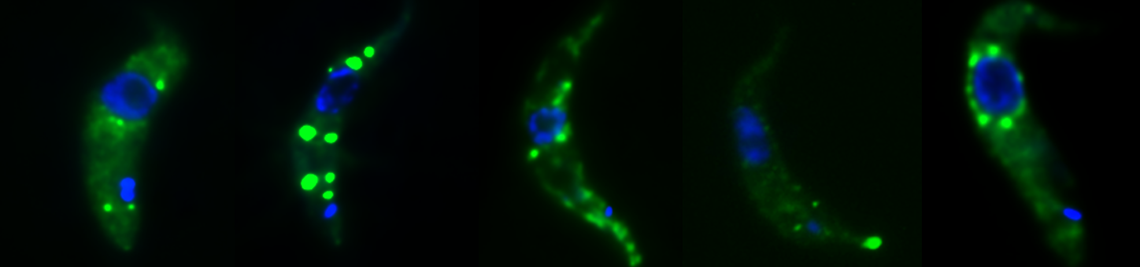
RNA granules are aggregates of protein and RNA. They are phase-separated from the rest of the cytoplasm and thus can be considered as organelles. There are several different types of RNA granules. P-bodies (processing bodies) are constitutively present and contain enzymes involved in mRNA decay. Stress granules form in response to a variety of stresses. P-granules are involved in mRNA storage during development. Neuronal granules are important for RNA transport. Even though RNA granules are a major focus of current research, surprisingly little is known about granule function and regulation. Granule purification methods were only recently developed and we have pioneered the purification of stress granules and trypanosome nuclear periphery granules. In the absence of transcriptional control, trypanosomes have an unusually large number of different RNA granule types, including some that are unique to the parasite. We aim to understand the function of the different RNA granule types in trypanosomes. In particular, we want to know how the formation and dissociation of the granules is regulated. We want to understand the mechanism of phase separation and we want to know, whether the granules are involved in the regulation of gene expression during the complex life cycle of the parasite. In particular the study of the trypanosome unique granules (nuclear periphery granules and posterior pole granules) will increase our understanding of the unusual RNA biology of the parasite.
Publications
Fritz M, Vanselow J, Sauer N, Lamer S, Goos C, Siegel TN, Subot I, Schlosser A, Carrington M and Kramer S. 2015 Novel insights into RNP granules by employing the trypanosome’s microtubule skeleton as a molecular sieve.Nucleic Acids Res43:8013–8032.
Kramer S. 2014. RNA in development: how ribonucleoprotein granules regulate the life cycles of pathogenic protozoa.Wiley interdisciplinary reviews RNA 5: 263-284.
Krüger T, Hofweber M, Kramer S. 2013. SCD6 induces ribonucleoprotein granule formation in trypanosomes in a translation-independent manner, regulated by its Lsm and RGG domains.Mol Biol Cell 24: 2098-2111.
Kramer S, Bannerman-Chukualim B, Ellis L, Boulden EA, Kelly S, Field MC, Carrington M. 2013. Differential localization of the two T. brucei poly(A) binding proteins to the nucleus and RNP granules suggests binding to distinct mRNA pools.PLoS One 8: e54004.
Kramer S, Marnef A, Standart N, Carrington M. 2012. Inhibition of mRNA maturation in trypanosomes causes the formation of novel foci at the nuclear periphery containing cytoplasmic regulators of mRNA fate. J Cell Sci 125: 2896-2909.
Fernandez-Moya SM, Garcia-Perez A, Kramer S, Carrington M, Estevez AM. 2012. Alterations in DRBD3 Ribonucleoprotein Complexes in Response to Stress in Trypanosoma brucei.PLoS One 7: e48870.
Kramer S, Queiroz R, Ellis L, Hoheisel JD, Clayton C, Carrington M. 2010. The RNA helicase DHH1 is central to the correct expression of many developmentally regulated mRNAs in trypanosomes.J Cell Sci 123: 699-711.
Kramer S, Queiroz R, Ellis L, Webb H, Hoheisel JD, Clayton C, Carrington M. 2008. Heat shock causes a decrease in polysomes and the appearance of stress granules in trypanosomes independently of eIF2(alpha) phosphorylation at Thr169.J Cell Sci 121: 3002-3014.